
Figments of Reality |
![]()
When we think of an amoeba,a fox ,an oak tree or a dinosaur,we think of them as part of nature.The amoeba fiddles about putting out pseudopods and ingesting food particles ,and that's about it. The fox runs through the bushes chasing a rabbit for dinner,and when it encounters the occasional bunch of subhumans on horseback it's too busy running from the dogs to debate the morality of bloodsports.[p7]
Few of us doubt that we evolved from creatures that,like most animals related directly to the natural world and thereby avoided all of the social problems that occupy our every waking minute and even assail us in our dreams. This question is central to the issue that will shape our narrative.What was it about this particular lump of rock,in this particular spiral arm of the galaxy,that made us the way we are? How is it possible for inanimate matter to turn into complex creatures like us with their own inner worlds of mind and imagination? Given that it is possible,why did it happen? Why us? Some will ascribe it to a God and be satisfied:we have nothing to say to them. Some will ascribe it to inexorable consequences of the fundamental laws of physics,and be satisfied:we have nothing to say to them either. We do have something to say to those who find the answer incomplete,people who think that our presence on this planet and our curious mental abilities deserve to be explained rather than explained away. [p8]
The point is that if you approach the questions from different directions you may find yourself wanting different kinds of answer,just as "God" may satisfy a priest in search of virtuous living but not a programmer in search of virtual reality. We do not so much observe reality as put together our personal representation of it and drape that back on to our perceptions of the external world.This facility is moderated by intelligence - the ability to reason and solve problems which is not merely a structural feature of large brains with intricate networks of nerves.[p9]
The bodies of living creatures are made from millions of different molecules,all of which trace back to the nuclear reactions in the cores of stars. Literally "we are stardust",as Joni Mitchell sang about in Woodstock. Particles building into atoms,and atoms into molecules - these we can comprehend,they're just like bricks building into a house [Actually,atoms are very much NOT like building bricks in a house,but for the purposes of the authors that is a sufficient model in the context they are using - A good indication of what atoms are actually like is found in The Tao of Physics].But houses don't develop a will of their own,get up ,and walk away.Living creatures did,and that's a real puzzle.How did inanimate,inorganic chemistry somehow generate the rich complexity of life Not all at once that's for sure. There was no wondrous special moment,pregnant with significance,at which life suddenly appeared on the planet. Instead,life emerged gradually from non-life.In this respect the origins of life are a bit like the origins of a person's life.There was a time when Maureen didn't exist.At what time did the egg,embryo,fetus,child become Maureen? At what time did it become human? Surely there was not a specific moment of becoming Maureen. A person is like a painting or a novel: it progressively comes into being. Maureen started with not-Maureen and gradually became Maureen.So it was with the origins of life. [p10-11] Carrying out a function is quite different from having a purpose. Molecular machines do not carry out functions because they want to do so :they carry them out because this is how they are made.Indeed,it is possible for them not to carry out their functions.In the same way,a rock carries out the function of rolling down a hill because it is suitably rounded and has significant enough mass for gravity to latch on to.But it does not have that rounded shape for the purpose of rolling down a hill. We mention this because human beings seem to have an innate tendency to confuse functions with purposes -so that,for example, "the sun keeps us warm" becomes "the sun was placed in the sky in order to keep us warm". This kind of purpose-centred thinking can easily lead to people worshipping the sun-god,not realising that the sun can perform the function of keeping them warm without either wishing to do so - or requiring worship to continue doing it.[p12-13]
Life seems very different from inorganic matter - it can move of its own volition,reproduce itself,consume other substances,respond to its environment.It is therefore hardly surprising that some people think that living material is simply a different kind of stuff from non-living matter. This belief is known as vitalism. Its greatest defect is that there is no evidence in its favour: none of this different kind of stuff has ever been isolated.If you take a living organism to bits,right down to the molecular level,all you find is ordinary matter [Recent innovations now show that the "more than the sum of the parts" that many people feel instinctively is the case is actually so,but there is no ghost in the machine - COMPLEXITY makes us a consequence of our parts,but we are not just what our parts make us,for more information on this,connect to the LINKS about Holism]. We humans are made from the same atoms as the rocks,water,and air around us. The inevitable conclusion is not that the ingredients differ:it is how they are organised.A living creature can be killed by bashing its head with a rock :it is hard to see how such a crass act can devitalise its esoteric immaterial substance,but easy to see how it can wreck its organisation. In the same manner a car is made from the same atoms as the sheets of polymer metal,sacks of aluminium powder,and cans of polymer from which it is assembled.Its ability to move does not arise because it is made from a different kind of matter:it is merely a consequence of how that matter acts when it is put together in a particular manner. An automotive engineer would be able to explain,in more than enough detail to send any partygoer in search of the drinks tray,what is involved in this organisation.But nobody ever made a car by going out and looking for a new kind of matter that has the ability to move when petrol is poured into it. There is a danger with the 'car' analogy if it is pushed too far. To some people, organisation implies the existence of an organiser, as the existence of a watch implies that of a watchmaker. This is a seductive line of argument, but there is no compelling reason to accept it. One of the most remarkable features of organic matter - and, we now realise, inorganic matter too under suitable circumstances - is its ability to organise itself .So in some ways a better analogy than a car would be a whirlpool, a tornado, or a flame: an organised structure that comes into being without conscious intervention. Our intuition is upset by self-organisation, probably because we seldom experience such behaviour directly: in our everyday world the only way to produce organisation is to work pretty damned hard to make it come about. Nevertheless, we are surrounded by and made from matter that is highly organised, and it must have got that way by some route. Either it has been organised by an organism-maker, or it has organised itself. The problems with the 'organism-maker' hypothesis have been rehearsed by philosophers and theologians for as long as anyone cares to remember. Its obvious advantages (it 'solves' the problem to many people's satisfaction) are countered by its equally obvious defects. For instance, who or what organised the organiser? And where is the organiser? The 'self-organisation' hypothesis has far more to offer to those who share the scientist's wish to understand nature and not just postulate it. It is a daring hypothesis, which does not solve the problem unless we can explain how and why living matter self-organises. It is becoming clear that there is nothing inherently self-contradictory in the idea that organisation sometimes comes 'for free', and it is also becoming clear that limited laboratory-scale systems and computer simulations indulge in self-organised behaviour far more often than we might have anticipated. Why, we are still unsure, but we know that it is so. Perhaps our universe is special in being like that; perhaps all universes must be, which.Which we don't know. The self-organising ability of life becomes clear only over long time-scales: compare an organism such as a mouse, today, to a lump of rock four billion years ago. One of the most obvious 'unusual' features of life, however, can be seen on far shorter timescales: its ability to reproduce. Life makes new life -and pretty much the same life. People make new people, cats make new cats, nematode worms make new nematode worms, and amoebas make new amoebas. This is an amazing ability, and it certainly looks very different from ordinary chemistry. However, we tend to underestimate what 'unaided' chemistry is capable of, and that distorts our assessment of how amazing or unlikely life is. Thirty years ago, biology was thought to be very complex and chemistry relatively simple. The chemical story of the origins of life seemed to require the construction of a conceptual pyramid of ever complicating processes, rising from the lowly plains of test-tube chemistry to the lofty heights of biology. Nowadays we understand that this picture is wrong. 'Unaided' chemistry - chemistry that does not require a living organism to make it happen - goes all the way up. Even simple unaided chemistry is a lot more complicated than the textbooks would have us believe. For example, if a mixture of two parts hydrogen to one part oxygen is ignited, then it explodes, giving water. The old textbooks see this as a single chemical reaction: 2H2 + O2 ® 2H2O. (We don't write this in the apparently simpler form H2 +O2 ® H2O, by the way, because reactions are about molecules, and a molecule of oxygen is O2 not O.) Newer textbooks will tell you that there are at least ten other molecules involved as intermediaries, and the more closely you look, the more of them you will find. The old textbooks tell you what to start with and what it ends up as, but not what happens in between. when reactions as basic as this turn out to be so complex, it is not surprising that more sophisticated kinds of chemistry are far more complex. Moreover, as our understanding of the complexity of chemistry grew, we also came to recognise that biochemistry is a lot closer to 'unaided' chemistry than we used to think. In fact modern industrial processes, which make extensive use of catalysts, sit right at the junction of 'unaided' chemistry and very similar biochemistry. Another reason why we are so puzzled by life arising from 'mere' chemistry is that it is very difficult to find, on the Earth, now, the kind of chemistry that long ago gave rise to life. This is because life has invaded all of the possible habitats for such chemistry, from the deep oceans, tens of miles deep in granite cracks, to high in the atmosphere - so their chemistry has been changed out of all recognition. Rusting would be a good example, except that on Earth it is nearly always 'assisted' by bacteria, who take a tithe of the energy. So let's imagine iron rusting on the surface of a lifeless planet. Recall the concept of catalysis: a molecule is a catalyst if it assists in the production of another molecule, or molecules, without itself being used up in the process. Sterile rusting proceeds by auto-catalysis - given a bit of rust on iron it catalyses more of itself .Such a process is recursive, it pulls itself up by its own bootstraps, so you need a bit of the product to get it started. (Stop worrying: we never said that that initial bit of product was produced by the same recursive process. See later) They are not, and neither is life. Thanks to the epic researches of Maurice Wilkes,
Rosalind Franklin, Francis Crick, and James Watson in the 1950S,
we know that one remarkable molecule - more properly a family of
very similar molecules - underlies almost all terrestrial life.
That molecule is DNA,
whose initials stand for 'deoxyribosenucleic acid' according to
taste)[p12-15].
DNA forms the genetic material of almost all organisms. A few viruses use RNA, 'ribose nucleic acid' or 'ribonucleic acid', but DNA and RNA come from the same molecular stable. DNA has a simple but clever molecular structure in which twin strands spiral like a staircase. The treads are made from four types of molecule called 'bases', held together by a framework of sugars and phosphates. This structure allows DNA to do two important things: encode information, and replicate. The information is represented by the sequence of bases, and includes such things as the structure of key proteins without which organisms cannot be built, and sequences that determine when they will be built. DNA replicates by separating the two strands, in which the bases are complementary to each other, and recreating a matching strand for each, thereby producing two copies of the genetic information from one original. (This description, though standard, is an over-simplification, but it is sufficiently accurate for our present purposes.) Throughout Figments we shall distinguish replication, the creation of exact or nominally exact copies, from reproduction, the creation of similar copies - in particular, similar enough that they too can reproduce. Normally DNA replicates, but when the occasional inevitable copying error - the technical term is mutation- creeps in, then the molecule is better thought of as reproducing. Although it is often described as such, DNA is not a self-replicating molecule: leave a mass of DNA in a beaker and you won't get more of it. It replicates only with the aid of many other molecules, known by names like transfer RNA, messenger RNA, and enzymes. We mention these merely to drive home that DNA needs an entire 'support team' in order to replicate: it no more makes copies of itself than a document in a photocopier makes copies of itself. Moreover, the fact that DNA contains 'information' is far less important than the physical (that is, chemical!) form that the information takes. All molecules 'contain' information- the positions of their atoms, for example, are a kind of information, as you will quickly discover if you build molecular models. The information in DNA is useful not because it is information, but because it is information stored in a form that other chemical machines can manipulate. As an analogy, the positions of the wood fibres that make up this page encode a huge amount of information, but when you read the page the only useful information - for you - comes from the letters printed on it. The process that allows DNA to replicate is another autocatalytic recursive cycle, only here it is a collection of molecules that catalyses itself. The DNA contains the defining information for the molecules in the support team. The support team helps DNA to replicate, and the DNA helps to replicate its own support team. Recursion often feels disturbing, but how else could a replicative process work? What makes recursive processes disturbing is the feeling that they can never get started - the 'chicken and egg' problem. Actually that's not a serious problem at all, just a case of sloppy thinking caused by incorrectly extrapolating the process backwards. It's relatively easy to get a replicative process started. What you can't do - without destroying the process - is stop it. The way to start a chicken-and-egg process is to create a suitable start-up configuration, one that is part of the process only the first time round. For example a non-chicken might be persuaded to lay an egg that grows into a chicken, whose eggs also grow into chickens, and so on forever. Clearly you can't play this trick if you start with a perfectly replicating non-chicken and absolutely nothing untoward happens to its egg, but if it is a reproducing non-chicken, subject to variations that do not affect the reproductive abilities of its offspring. there's no conceptual problem at all - just a technical one of actually making the trick work. The answer to the hoary philosophical teaser then becomes no more than a question of definition. Is a chicken egg one that was laid by a chicken, or is it an egg that grows into a chicken? In the former case. the chicken came first (from a non-chicken egg), in the latter case, the egg came first (laid by a non-chicken). There are other ways to get a replicative or reproductive system started. One is for it to 'piggyback on a pre-existing replicative or reproductive system. This is how documents replicate: they piggyback on photocopiers, which are replicated by humans working in factories. The photocopiers in turn piggyback on human reproduction. Of course it's not possible for every replicative/reproductive process to piggyback on a previous one. or else there is a genuine chicken-and-egg problem. so at least one process has to get started some other way (and act as a start-up configuration for everything that subsequently piggybacks on it). That other way is best described as 'scaffolding': before the replicative loop closes up, the process is assisted by something else, which drops out of the loop permanently after it is closed. Once a system acquires the ability to replicate. it spreads rapidly and takes over any disorganised substrate. Although the loop formed by DNA and its support team is in principle replicative. In practice it is 'only' reproductive. The procedure is so complex that it seldom takes place without errors. Moreover,in sexually reproducing organisms, the reproductive procedure introduces 'mix-and-match' modifications. This should not be thought of as a defect. Reproductive systems are much more interesting than mere replicative ones, precisely because they can change. Replication is just the same thing repeated forever. Reproduction has room for flexibility - it can produce a chicken from a non-chicken's egg. [p17-18]
Burgess Shale - evidence of evolution.Why science is not another belief system. The realisation that most scientific observations are context dependent,has led some philosophers to argue that science is a social construct which has nothing to do with reality and is solely a matter of human convention. This argument stems from the entirely sensible modern perception that scientific 'truth' is not absolute, but depends upon having some agreed common conceptual framework. However, the belief that science is solely a construct, which by implication could be whatever scientists decided to agree upon, is really very silly - however elegantly it may be phrased - because it ignores a very important aspect of these conceptual frameworks. They are not arbitrary: they are the outcome of a previous scientific process. For example, scientists cannot make objects float skywards merely by agreeing among themselves that the force of gravity acts up rather than down. There has to be a reality check. Science has more stringent reality checks than any other area of human activity, and applies them more frequently. Religion hinges upon faith, politics hinges upon who can tell the most convincing lies or maybe just shout the loudest, but science hinges upon whether its conclusions resemble what actually happens. Not so long ago we might have said 'whether its conclusions are true', but the idea of science as absolute truth has gone the way of the dodo. Because human beings experience reality indirectly, through the medium of their senses, there is room for genuine and reasonable disagreement about the nature of reality. Even a reality check must have a contextual element. At any rate, the role of science is not to establish some kind of factual data-bank about nature, but to help us understand nature. There are many different degrees of understanding, and many different kinds of understanding. Some aspect of nature may be not understood, or slightly understood, or fairly well understood - although it is unlikely that it will be fully understood. More subtly, it may be well understood from one point of view but an impenetrable mystery from another. Thus we may understand that a flower petal is red because it contains a particular pigment, but fail to understand why the flower uses that pigment and not a different one.[p36]
4 Winning ways A species of viperine snake, which is not poisonous, has evolved three ways to protect itself against predators. The first is camouflage, so that it gets 'lost' against its background. However, its camouflage is very similar to that of the poisonous adder, which leads to the second method: mimicry. If a predator sees through its camouflage, it exploits the resemblance to an adder by behaving like an adder. But if this doesn't work either, for example when the predator is a crow, which kills adders, it adopts the third strategy. It flips about like a demented rope, and then it arranges itself on the ground to look for all the world like a dead snake, lying on its back in the dust at an awkward angle, with a vaguely bloated look... However, if it is now turned on to its front, it promptly and energetically flings itself back into its 'dead snake' pose. The background theory and philosophy is now out of the way, and we are ready to begin the journey from molecules to minds. It is a journey which, at every stage, involves the concept of evolution. Evolution is a general mechanism whereby systems can 'spontaneously' become more complex, more organised, more startling in their abilities. So in this chapter we shall take a closer look at the evolution of living creatures, paying especial regard to the idea that evolution can profitably be viewed as a game - indeed a nightmare game, according to the classification of Chapter 2. Evolution plays games with the forms and behaviours of organisms, and also with human sensibilities. In a book of the same name, Daniel Dennett calls evolution 'Darwin's dangerous idea'. The concept of evolution is dangerous because it provides a compelling rebuttal of the central tenet of nearly all religions: that the world we live in was created by a supernatural being. Life must have been created, the argument goes: it is too complex and structured ever to have arisen through the normal workings of ordinary matter .Not so, says Darwin: there are excellent reasons to suppose that life evolved of its own accord from more simply organised matter.Just as, centuries earlier Galileo's astronomical observations threatened to upset a theological applecart based upon the Earth as the centre of the universe, so Darwin's deceptively simple idea threatened the keystone of nineteenth century theology. Darwin's idea is not only dangerous, in this cultural sense: it is subtle. In this chapter we shall tell the story of evolution in at least two versions: the standard textbook version, which focuses on genes, and a contextual version, which focuses upon emergent dynamics in phase space. Our treatment of the latter viewpoint will be on a fairly general level, though we will invoke Darwin's finches to provide some more concrete mental imagery. In the next chapter we will move from generalities towards specifics, and in Chapter 6 we will derive a complementary viewpoint to genes, based upon the interactions of organisms -in particular the concept of privilege, in which parents provide their children with a head start in life. We outline these approaches now, and develop them in greater depth as the book proceeds. The textbook gene-based version goes something like this. All animals produce too many offspring, so that there is no room for all of them to breed. In consequence those offspring that do manage to breed contribute to the next generation, while the others do not. Some attributes assist in the process of breeding, or surviving until breeding age - strength, speedy reactions, the ability to be as quiet as a mouse when the cat is nearby, whatever Suppose that one animal - call him Fred - manages to breed. If the attributes that allowed Fred to breed are inherited by Fred's offspring, then they will possess the same advantage. This doesn't mean that they will necessarily get to breed, but it does mean that their chances of producing Fred Jr. are improved. Over several generations this slight advantage is likely to lead to a preponderance of Freds, with his special attributes. The population has now changed: nature has winnowed the creatures that might have been produced and chosen mostly Freds. This process is known as 'natural selection', and it ensures that the resulting creatures have 'good genes'. This is the essence of the story as it is usually told, with a few extras like the necessity for heritable differences and bells and whistles like dominant/recessive alleles (alternative versions, such as brown or blue eyes, one of them taking precedence) of each gene. The 'emergent dynamic' version places the emphasis on organisms rather than genes, and sees the textbook process as a complicated way of describing something much simpler: a dynamic on phase space. In this view, organisms change because the geography of the surrounding space-of-the-possible makes change inevitable. Evolution runs 'downhill' in its phase space. The hidden complication is that the dynamic is emergent, not prescribed once and for all, and the phase space changes complicitly with it. This organism-based view places emphasis on many different things, among them privilege. We shall go into this idea in depth in Chapter 6, but a short preview will help set the scene now. Instead of concentrating on organisms competing for how many progeny they pass their genes to, this alternative theory focuses attention upon how the parents provide their offspring with a head start, and how siblings compete for the privilege of growing up. If you are a baby starling, then your immediate source of food is your parents, so that the most immediate short-term competition comes not from prowling cats, but from your brothers and sisters. Darwin saw that this kind of 'sibling rivalry' must be very intense, and for him it was the major driving force of evolution. Of course it is important to be able to avoid the prowling cat too, but you have to grow up first, and that won't happen ifyou don't get a fair share of the food. The way adult birds feed their young is grossly unfair: the one who is pushiest, makes the most noise, opens the biggest gaping beak, gets most of the food. 'To him that hath shall be given.' Unfair it may be, but this simple strategy has survival value: it ensures that at least one chick gets enough nourishment to reach the stage of leaving the nest and foraging for itself.Divide your food supply among too many young, and if it is inadequate - as it often is - then they all die. An extreme instance occurs in the fish eagle, which typically starts by trying to bring up three chicks. What you see in the nest is one large chick, one medium, and one small, but that's because they hatched at different times. If there is too little food, which happens in most years, the large and medium chicks eat the small one; if necessary the large chick then eats the medium one. It is conjectured - but not yet established - that in very bad years the parents end up eating the big chick. This approach violates many of our cherished sensibilities, but it makes perfectly good sense to the fish eagles, who in effect use their chicks as a living refrigerator to keep food in usable form, and can thus adopt a strategy that avoids putting needless effort into a baby that won't become a breeder. Those are the alternative positions that we shall compare, contrast -and eventually combine. We start by taking a closer look at the textbook story of 'good genes'. In order for competition - either between species or within them - to affect future generations, some kind of hereditary factor must pass from one generation to the next. The system must have some kind of 'memory' of what works, and what does not. Darwin knew that such a hereditary factor must exist, but he didn't know what it was. Nowadays we know that the basic genetic material is DNA, and we say that the breeders - the animals that survive to produce the next generation - are most 'genetically fit'. As we've just said, alternative possibilities for a gene - more properly, genetic differences - are called 'alleles'. Natural selection favours some alleles over others, so textbooks of population genetics take a lot of space describing how to calculate the relative genetic fitness of different alleles. A particular allele may be fitter if creatures that possess it are more able to survive than if they possess any of the alternative alleles. The simplest way that this can occur is if the alleles correspond to clear-cut 'characters' - features of the organism such as shape, behaviour, or colour - and particular characters enhance survival value in particular circumstances. However, despite a widespread belief to the contrary, most alleles do not correspond directly to characters. Alleles may also be fitter if they enable the organisms that carry them to increase the representation of those alleles in the next generation. This is not quite the same thing as helping the organisms themselves to survive, though the distinction often doesn't matter, because if you don't survive to breed then your fitness is zero. However, characters needed for survival often mitigate against breeding: for example in order to survive you must not be visible to a predator, but in order to breed you must be obvious to your intended mate. Again, staying away from your own species is a good survival trick, because they are the most likely source of parasites and disease; but if you belong to a sexual species that strategy is hopelessly poor when it comes to breeding. Organisms have evolved special tricks, such as elaborate mating rituals, to cope with these contradictory tendencies. Some evolutionary biologists, the best-known being Richard Dawkins, tell us that DNA is in the driving seat of evolution. This view is known as 'neoDarwinism': it is Darwin's original idea of natural selection, together with a specific statement of what the 'unit of evolution' is. The neo-Darwinists' stories have become increasingly subtle over the years: for example they now say that it is not just your DNA that improves its chances of survival by making you better.Alleles can sometimes increase their chances of appearing in the next generation by promoting the survival of close relatives rather than that of the actual breeder.Worker bees, which do not breed at all, provide one of the best known examples. This effect leads to the concept of 'inclusive fitness' of an allele: when calculating an allele's fitness, you must take account of all relevant copies of that allele, not just those in one particular organism. In Chapter 1 we mentioned that DNA has a fascinating structure, the famous 'double helix', composed of two intertwining strands made from four types of 'base'. The bases rejoice in the names cytosine, guanine, thymine, and adenine, normally abbreviated to C, G, T, A. [p77-80]
The finely tuned machine We can now go further, and consider the question of evolution on a deeper level. Evolution strikes many people as being highly implausible, because today's world is so amazingly complicated and finely tuned. They find it hard to see how such a wonderful, interlocking 'machine' could have come about through the blind workings of chance. What they often fail to bear in mind is that today's world is a snapshot of a game that has been in progress for five billion years. If it were possible to watch the entire game, the current position would make much more sense. We can't do that, but we can make reasonable inferences based upon our understanding of chemistry, biology, geology, the fossil record, and anything else that seems to be implicated in the game. It is a game that began when the first replicating molecular network accidentally - but if Kauffman is right, inevitably - assembled, and it has never stopped. It is the game of Survival, and you can consider it as having either countless trillions of players (all living creatures) or only one (the global ecosystem). Our language lacks the words, and our science lacks the concepts, to describe this game adequately, so we give it a label: Evolution. Putting a label on a concept doesn't mean that you understand it, but at least it makes you recognise that there's something to understand.[p95]
The Galapagos finches are not anathema to Evolution One result, which we can see clearly from our 'game' analogy, is that the survival strategy becomes increasingly ritualised. This point deserves examination at length, so that's what we'll do now. When we look at an ecology - an interacting system of organisms, animals and plants and insects and worms, or whatever - we do not see the Game of Evolution being played out in the way our liar-to-children teaching myths seem to suggest, with badly designed animals locked in mortal combat with their eventually triumphant superiors. Part of the reason is that the real story is more subtle anyway, but in fact the game is being played, still - but by professionals rather than amateurs. The winning strategies have become entrenched, and most moves have become routine. Organisms still experiment with novel moves, but hardly any of those succeed, and nearly all of the failures are removed when still very young, so we don't notice them. Think of the millions of acorns that never get to grow up into oak trees. The ecosystem has become 'canalised', just like the genetics of wild-type organisms; that is, it is much more able to survive environmental changes without (apparently) altering its behaviour. This is an important point, affecting our entire conception of how evolution and ecology are related, and it is worth spending enough time to appreciate it. As an example consider 'Darwin's finches' on the Galapagos islands. There are thirteen distinct species - some rest in trees, some rest on the ground, some inhabit cacti, some live amid the mangroves, some eat seeds, some eat snails, and so on. Between them they have occupied the main 'ecological niches' open to birdlife in this very impoverished ecosystem. The current division of resources is stable, in the sense that new species do not seem to be evolving, although we can detect evolution in microcosm as the proportions of different species vary from year to year in response to changes in environmental conditions. In what sense, however, are they really distinct species? There are two accepted concepts of 'species': one is taxonomic, the other reproductive, and they do not always agree. Taxonomically two (sexual) organisms belong to the same species if they resemble each other in sufficiently many ways, including being able - potentially - to interbreed, giving rise to fertile descendants. The reproductive definition, promoted energetically by Ernst Mayr, is that a species is a breeding group. If organism A can in principle interbreed with organism B to yield fertile descendants, then A and B belong to the same species. Now, taxonomically, the thirteen types of Darwin's finches are separate species. Reproductively, however, they are not: they are thirteen 'morphs' within a single species. Birds from different morphs can interbreed with each other to produce fertile 'hybrids', cross-bred individuals whose genes combine those of both morphs. For example a snail-eating finch can interbreed with a cactus finch. Nevertheless, despite this 'gene flow', the morphs remain separate. Why? There are at least two possible reasons. One is that since different morphs prefer different habitats, the opportunity for crossbreeding is small. The cactus finch likes the dry parts of the island and seldom gets an opportunity to breed with a snaileater that stays in the wet parts where the snails live. However, this cannot be the full explanation, because there is plenty of overlap at the edges of habitats. Over a relatively small number of generations, the hybrid gene combinations would diffuse widely. However, you don't find them. The real reason is the Grim Sower. Hybrids do occur but they do not survive in the current ecology of the Galapagos islands. A finch that combines some of the strategies of a snaileater with some of those from a cactus finch falls between two stools, and cannot compete well with either It searches for damp-loving snails among the dry cacti, perhaps. So the hybrids die young, and do not go on to breed. There is nothing 'wrong' with the hybrids. All of their genes are 'good' - inasmuch as this concept makes sense - because they come from 'good' parents. But the combination - not so much of genes but of characters - doesn't work when the thirteen existing species have already carved up the territory to suit themselves. This view of how organisms partition their environment into 'eco-territories' shows that we used the wrong word when we talked of species 'occupying' niches. Actually they create niches through their collective and complicit interactions with each other and the rest of their environment. After the ecosystem has 'settled down', the viable niches are pretty well determined and segregated. The winners are the professional snaileaters and the professional cactus-eaters, whose strategies have a proven track record. The hybrids that nest among the dry cacti, but want to eat the moisture-loving snails that inhabit the marshes ten kilometres away, are incompetent amateurs, making the wrong moves; and they lose every game they play. The Grim Sower ensures that they go on playing long after their incompetence has been amply demonstrated - and they go on losing.
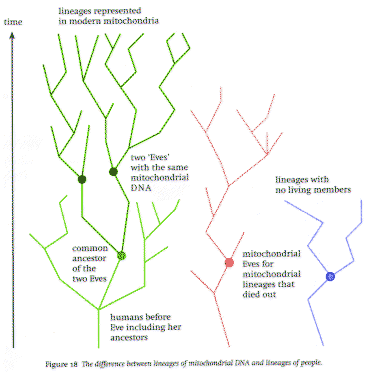
Why mitochondria does not support the biblical EVE At last we are equipped to follow the story of mitochondrial Eve,erstwhile universal mother. The 'Eve' metaphor trips off the tongue of anyone brought up in an even nominally Christian country. Too easily perhaps? Was there genuinely a single 'Eve' - one female ancestor of all of today's humans - or were there several? This is a loaded question, as the 'Eve' terminology reveals: it bears upon the authenticity of the Christian creation myth. So it was not a great surprise that in 1986 the news of scientific proof that all of humanity did indeed descend from one female ancestor attracted a lot of media attention - predictably greatest in the United States, where it came as welcome news to the Bible Belt and its gaggle of 'creation scientists'. 'Mitochondrial Eve,' she was named, because the evidence for her existence came from an analysis of resemblances between DNA sequences in the mitochondria of human cells. She lived in Africa between 100,000 and 200,000 years ago. The evidence was quite compelling, based on tracking such resemblances backwards in a molecular family tree, as just explained. The scientists analysed large numbers of sequences derived from modern human mitochondria, from people all over the world, and tracked their DNA genealogies backwards. The evidence seemed to point back to a single human being as the Mother of Us All. What exactly was that evidence? The human genome contains some 6 billion DNA base pairs, most of it inside the nucleus incorporated into chromosomes: half of it in each set of 23 chromosomes inherited from each parent. However, about sixteen thousand base pairs of DNA exist inside mitochondria, which are organelles of the cell outside its nucleus. This is a short enough sequence to be handled easily when tracking differences that have accumulated over time through mutations. Mitochondrial DNA has a distinctive pattern of inheritance: it always comes from the mother Analysis of mitochondrial DNA from more than a hundred ethnically diverse modern individuals shows that it all tracks back to a single DNA sequence, 'coalescing' some 200,000 years ago. Conclusion: we all have the same great-great- ...-great-grandmother, with about ten thousand greats (at twenty years per generation, say). That's Eve. Only recently has it been shown that this entire story is based upon a misunderstanding. Almost predictably it arose from a conceptual confusion between the genealogies of genes and those of individuals - yet another mistake brought about by taking the 'DNA-as-blueprint' image too literally (A genealogy is a 'family tree', a list of predecessors going backwards into the past.) This work, summarised in the journal Science by Francisco Ayala in December 1995, tests the theory of mitochondrial Eve by playing the same game with certain genes from within the nucleus, known as DRB1 genes. They are involved in the immune system. A typical DRB1gene contains 270 base pairs, and is thus easily sequenced in full. DRB1 genes are highly polymorphic in humans, meaning that different people have different DRB1 genes: there are 59 distinct morphs - 59 choices of DNA sequence. There is strong evidence that human DRB1 genes go back a very long way indeed. For example the human DRB1gene known (imaginatively) as HS*1103 resembles the chimpanzee DRB1gene Pt*0309 more closely than it resembles another human DRB1 gene, HS*0302. Therefore the two human genes diverged from each other earlier than the chimpanzee and human lineages diverged from each other. (There is no contradiction in this, even though we call them 'human' genes. They are in humans now - but they were in the common ancestor of humans and chimpanzees in the past.) By studying the rate at which mutations normally occur, it can be inferred that these two human genes diverged about 6 million years ago; the divergence between human and chimpanzee lineages occurred soon after For comparison, the orang-utan lineage diverged from that of the chimp-human ancestor about 15 million years ago, and - as we have just stated -H. erectus emerged about 1.7 million years ago. These times are estimated by the same method that produced the 200,000 year estimate for Mitochondrial Eve - if you dispute that method, fine, but then Eve goes out of the window straight away The genealogy of the 59 different human DRB1 genes coalesces about 60 million years ago - which, coincidentally or not, is about the time of the great mammalian radiation brought about by the demise of the dinosaurs. The same kinds of estimate show that 6 million years ago about 32 different members of today's human DRB1 gene lineages were still in existence. This implies that at least sixteen distinct human ancestors existed then, because each human can possess at most two DRB1alleles, one in each set of chromosomes. The same reasoning shows that there must have been at least sixteen ancestors in the human lineage after it diverged from that of the chimpanzees, so Eve must have had, at the very least, fifteen Adams to keep her company Actually a more careful mathematical analysis shows that throughout the last 60 million years human ancestral populations have generally contained at least a hundred thousand individuals, though there is a slight chance that they occasionally may have shrunk to about ten thousand. The theory of one Eve and 99,999 Adams really won't wash - she wouldn't have time to make enough babies - and it is much more likely that there were about 50,000 of each. So why does our mitochondrial DNA all go back to one sequence? Possibly there did exist a Mitochondrial Eve, but she is not the Mother of Us All: she represents a particular molecular sequence for mitochondrial DNA, embodied in a population of women possessing that molecule, from whom all modern mitochondrial DNA molecules descend. We got a lot of our other DNA from other ancestors in any case, and all we can infer is that 200,000 years ago all of our 50,000 or more female ancestors happened to have the same mitochondrial DNA. Of course, if we push Mitochondrial Eve back sixty million years, to long before she was human .. But then she sure as heck wasn't created in the same image of God that we are said to embody Perhaps the most ironic aspect of this story, apart from the credulity with which the original results were received, is that the analysis of DRB1 genes implies a human ancestry stretching back some 54 million years before humans appeared on Earth. Far from supporting the creation story, Mitochondrial Eve supports evolution. We have told this story from the conventional point of view about convergence of alleles and molecular clocks, and many biologists are happy with that viewpoint. We're not, and it's worth taking a few moments to indicate some of our objections. One (which is another objection to the interpretation of' 'mitochondrial Eve' that does not require games with DRB1 genes) is that the analysis applies only to those women who have descendants alive today. It does not tell us anything - by its nature cannot possibly tell us anything - about other women who were alive at the same time as 'Eve', but whose lineages died out. However, in real populations lineages die out all the time: the most familiar analogy is the disappearance of surnames, so that as the centuries pass we find more and more Smiths but after a while no Chaucers (well, not in the Coventry phone book, at least). The Biblical Eve is supposed to be not just the first female ancestor of everybody alive today but of everybody alive ever. And that you cannot deduce from modern mitochondrial DNA (see figure 18). The next objection is methodological. By starting with a sample of only a hundred ethnically diverse individuals, you run a very real risk of omitting many rare morphs that occur somewhere in the human population - but very infrequently. As an analogy imagine that you are surveying patterns of human wealth by choosing a hundred financially diverse individuals, at random . You will without doubt find many instances of grinding poverty, because huge numbers of people are very poor, but even in a developed country like the UK you may well miss all of the millionaires (there are less than a hundred thousand in a population of over fifty million). You will almost certainly miss the billionaires -just as the rare bloodgroups like 'Kell', occurring only in a few families, were missed by everybody until very recently At any rate, if additional morphs have to be factored into the coalescence calculations, it is likely that the estimated coalescence time will increase;and if there are lot of missing morphs,it will increase considerably. So what the analysis showed, at best, was that (say) 95% of people alive today had a female ancestor among the 'core' group that 200,000 years ago,all had the same mitochondrial DNA. That doesn't sound like Eve to us.
We begin with a topic that has greatly excited many philosophers: along with Dennett, we think it is a complete red herring. We mention it because you may well run into it, and deduce from the awe in which it seems to be held in some quarters that it is a serious contribution to the subject - which it is not. This is the concept of a 'zombie', which is exactly like a human in every respect except that it is not conscious. Zombies are thought experiments, often used to focus attention on the (alleged) ineffable quality of consciousness that is so different from the physicochemical activities of material brains. imagine a zombie, the argument goes. It functions just like a real human being, right down to the electrical activity of its brain cells - but you know it's not conscious, because that's how you imagine it. Conclusion: consciousness is different from the physical activity of a material brain. There are so many things wrong with this suggestion that it is hard to Know where to start. The main one is that it's a cheat. It's a Silly question in disguise, one of the 'but what if you saw a ghost?' variety, which we dismissed in Chapter 2. Suppose that mind is an emergent property of brains, as seems likely. Then mind is inaccessible to detailed reductionist analysis - not because brains contain some ineffable non-material quality, but because reductionist explanations of all emergent properties have to traverse Ant Country. Now in Ant Country, the chain of logic that links the emergent property to its reductionist causes exists; it is just far too intricate for anyone to comprehend. Nature, running trillions of causes-and effects in parallel, traverses Ant Country with ease - it is we who get bogged down. So if mind is an emergent property of the brain (in the context of a human being in a human culture, we must add) then it would not be possible to have a creature identical in every physical respect to a human being, without consciousness emerging from its structure and activity -just as it does in a human being. Emergence is not the absence of causality, something that disconnects effects from causes: it is causality too intricate to follow in detail, but which still achieves its effect. You might as well encourage people to imagine a zombike, which is exactly like a bicycle in every way except that it does not move when the pedals are pushed. Oh, mystic miracle of ineffable immateriality, the source of motion in a bicycle is not anything physical! Or a plant that has exactly the same molecular structure as grass, but is pink with blue spots: wow, colour is not a physical property at all! Or as Dennett suggests, imagine a creature that is exactly like a human being right down to its DNA structure, which walks and talks like a human being, and is not alive. In any case, if you really do want to allow that game, how about imagining a being that is exactly like a pocket calculator in every respect, except that it is conscious?
In order for a thought experiment to illuminate a problem, the experiment must bear some relation to reality. Being able to imagine a zombie, a ghost, or a conscious pocket calculator does not imply that such a thing can actually exist, so any 'ah, but what if one did exist?' argument falls flat on its face at step one. But such arguments are insidious: as Dennett rightly points out, they 'work' because they invite people not to use their imaginations. The question 'is consciousness an emergent property of material brains?' does focus our attention, and it is an excellent mental exercise to try to imagine how such a thing could possibly be. Notice that this question is not posed in a manner that rules out either of the answers 'yes' or 'no' in advance. In contrast, the zombie concept actually invites us to try very hard not to imagine how such a thing could possibly be: and if we accept the possibility of zombies then we immediately concede that consciousness is not a property of material brains, Thought experiments like this rely upon us limiting our imaginations, and in this respect the zombie resembles Searle's Chinese room, which plays a similar distracting role with regard to intelligence.
So much for zombies. Another common misconception
in the literature concerns the location of mind, or
indeed consciousness. where is it? In the brain? Sure, but where
in the brain? All over? No, that's not a location. But surely if
mind exists in a material sense, then it must be somewhere?
Sounds sensible - until you put your own mind in gear for a
moment. Mind is a process, not a thing. Processes take place
within things, but they do not have a specific location. where
in a car is its ability to move located? In the wheels? No,
because you need an engine too. In the engine, then? No, because
you need the wheels. In both (along with transmission and so
on)? But that's not a location!
That brings us to religion: in particular, the pervasive concept of God. Religion excites strong emotions even in non-believers: it creates cohesiveness within a single cultural group but is often responsible for deep divisions between different ones. Belief in gods goes back as far as recorded history, but modern religions have generally converged on belief systems centred around a single 'invisible' being, credited with a range of supernatural powers. We think this happened because cultured humans relate to the world in a new way, thanks to extelligence. Because we and most of our readers belong to the Judaeo-Christian tradition, we shall set our discussion in that mould, taking as an example the Old Testament God of Abraham. What we shall tell is pure conjecture, at best a myth or a parable, almost certainly wrong in many details; in particular we may well have given it the wrong protagonist. But we suspect it is at least 'meta-true', describing a general process that did occur. Abraham, we are told, was a shepherd. His culture - possibly that in the region of Ur of the Chaldees - surrounded him with a whole list of rules, recommendations, and prohibitions. Around him were other cultures, which he knew by the names of the gods: Marduk and Baal, Anat and El, represented by graven images' - masks, altars, temples ... the numinous appurtenances of many tribal peoples today. Abraham was unimpressed by such tawdry displays. Instead, he was awed by how much more 'something outside him' knew than he did. That something was the early blossoming of extelligence. It knew when to plant crops when to reap them, what to eat, how to run your life to ensure prosperity, even how to ensure that all of this was passed on to your children. Abraham could see that his own intelligence was tiny by comparison to that outside authority on his own life, invisible but immensely benevolent, knowledgeable, and successful. and not represented by a graven image. It was best described as That Which Is (the meaning of 'Jehovah'), the God with no name and no image. It is a very small step from 'There is Something out there' to 'There is a Being out there'. It is also a very small step from 'That Being looks after my affairs and makes sure that my sheep, wives and children prosper' to 'That Being regulates the lives of the sheep, and the wild animals, and all the patterns that I see in nature... Gosh, It probably makes the sun come up in the morning, and has put all of those amazing stars in the sky! Now, that is a Being worth worshipping; I'll make sure Isaac gets a load of this...' If we are right, then monotheism arose from a form of ontic dumping, in which extelligence was turned into a thing. We are not suggesting that modern Jews and Christians have retained this simple view, merely that perhaps it began that way. The concept of a single all-powerful, all-knowing, all-seeing God then became embedded into the very extelligence that spawned it. A Zarathustran visiting the Earth billions of years ago would not immediately see all the descendants - ammonites, octopuses, squids - implicit in a particular tiny flatworm with a shell; similarly it is not obvious that Abraham's conception would lead to the glory of Solomon, the Archbishop of Canterbury, or Mecca. An alternative way for
humans to relate to nature also grew from the new extelligence
that Abraham founded, but it grew along a different
branch:science. Religion keeps its snooker-break going because
it passes on its beliefs to succeeding generations, and
reinforces them with the same awe that led Abraham to the idea
in the first place. Science
does more than this: not just passing on the belief
that planetary orbits are ellipses, but passing on the need, and
the technique, to constantly test that belief against the real
world - and to test it by looking for evidence that it is wrong,
not just by selectively accumulating evidence in its favour. As
a result, science has been far more successful as a way of
understanding nature. Science and religion, then, may have had
very similar origins. Both are attempts to systematise
humanity's relation to its extelligence.
|
![]()
[From "Figments of Reality" by Ian Stewart and Jack Cohen]
|
Related Articles |
|
Ordinary Miracles Complexity Theory |
|
Everyone's a Winner Quantum Theory |
|
A Random Walk in Arithmetic by Greg Chaitin |
| Rationally Speaking Massimo Pigliucci's page |
|
Random Reality by Greg Chaitin |


 Inevitably
there is a male parallel to Mitochondrial Eve: ZFY Adam. A male
Inevitably
there is a male parallel to Mitochondrial Eve: ZFY Adam. A male
 IN THESE days, Big
Theories about
IN THESE days, Big
Theories about