What is an organism?:A discussion
Brian Goodwin, Biology Department, Open
University, Milton Keynes.
Richard Dawkins, Department of Zoology,
Oxford University, Oxford.
ABSTRACT
Two fundamentally different ways of describing organisms are opposed. One,
arising from embryology , describes organisms as irreducible wholes that
evolve not just through incremental change but through transformation. The
other, rooted in population biology, focuses on the genetic substitutions
that make evolutionary change possible.
INTRODUCTION
Biology is the study of life, and life comes in the form of organisms. One
might expect, then, to find in biology some generally agreed description
of what an organism is. However, two basically different descriptions of
organisms have arisen from different ways of understanding and interpreting
biological processes. One of these is directly descended from Darwin' s view
that organisms are the products of natural selection operating on hereditary
variations in reproducing populations. The modern version of this theory
was founded when Darwinism was combined with Weismann's doctrine, a theory
that separated the organism's somatoplasm from the hereditary determinants
that direct the development of the somatoplasm into an organism of a specific
kind. One participant to this discussion, Richard Dawkins, has extended these
ideas in a series of books entitled The Selfish Gene, The Extended
Phenotype, and The Blind Watchmaker. Here organisms are described
as vehicles for the genes that built them. A guiding metaphor of this work
has been the notion of the selfish gene, a unit of inheritance that co-operates
with other units of inheritance only because it has profited from their help
in constructing organisms that are mechanisms for its own perpetuation. This
co-operation arises from selection in favour of those genes that do well
against a background of the other genes that it encounters in the gene
pool.
A second view of organisms sees them as wholes that cannot be reduced to
the products of genes and the action of natural selection. Here organisms
are described as members of a class of complex dynamic system with distinctive
properties of order and form. Without an understanding of these special
characteristics of organisms it is not possible to make sense of the phenomena
of evolution. Organisms evolve not just through incremental change but through
transformation. This perspective, which belongs to a long tradition of biological
thought that is found in the work of biologists such as Waddington, Needham,
D' Arcy Thompson, and William Bateson, has recently been revived in Stuart
Kauffman's The Origins of Order (1993). It is here represented by
the other participant of this discussion, Brian Goodwin. On Goodwin' s
initiative, the two met for a public debate in London chaired by Patrick
Bateson in 1990. This paper grew out of that debate.
BRIAN GOODWIN. ORGANISMS ARE WHOLES AND EVOLUTION IS
TRANSFORMATION
Richard Dawkins' position is that organisms are survival machines constructed
by their genomes and adapted by natural selection to existence in a particular
habitat. Let me explain why an organism is not a machine; and then why it
is not constructed by its genes. In doing so I'll tell you what I believe
organisms are. What will then emerge is how the theory of natural selection
can be true and still not explain basic properties of organisms, such as
their form or morphology. Natural selection is a principle of dynamic stability
which certainly has significant use in biology. But as an explanatory principle
of biological phenomena it is limited to questions of persistence and says
nothing about the actual structure of organisms.
What is the difference between a mechanism and an organism?
I'm going to use a definition that goes back to the 18th century philosopher,
Immanuel Kant, because it focuses On a crucial
distinction that tends to get blurred in modern biology (see Cassirer, 1981,
for a discussion of Kant' s views). A mechanism, said Kant, is a functional
unity whose parts exist independent of one another. The clock was the
paradigmatic example of Kant' s time, and it continues to serve the same
purpose today, as in Dawkins' The Blind Watchmaker. An organism, on
the other hand, is a functional and a structural unity, in which the parts
exist for and by means of one another. That is, the parts of an organism,
such as eyes, brain, and limbs, or flowers, leaves and roots, do not pre-exist
before being assembled into a functioning unity, as in a machine. They are
produced by the activity of the organism itself, which is self- generating.
This activity is the process called development, the transformation of the
fertilized seed or egg into a functioning whole with a specific structure
that can be described in terms of particular arrangements of distinctive
parts. What modem biology has tried to do is to describe the organism in
terms of the activities of its genes, that is, in terms of the molecules
out of which it is made.
What I shall now show is that although organisms are made up of gene products,
knowing that these products are, where and when they are produced, doesn't
explain organisms. The fact is that composition rarely explains form. Knowing
what a liquid is made of doesn't explain its form, as when it flows in a
spiral vortex down a drain or forms waves under the action of air passing
over its surface. Even in crystals, composition doesn' t determine
form: carbon can exists in the
form of diamond or graphite. Knowing the composition of the planets certainly
doesn' t explain their elliptical motion round the
Sun. In general if we want to understand form either as structure or
activity we have to know more than composition. We have to know the principles
according to which they system is organized as expressed in, say , the equations
of fluid flow or of crystal formation or the dynamics of moving bodies. We
also have to know the conditions to which the system is exposed- Organisms
are no exception to these physical principles. If we want to describe their
most basic properties, such as how they are generated, we have to understand
the principles according to which they are organized. Knowing their molecular
composition may be very useful in helping to figure out these principles;
but describing organisms in terms of a catalogue of gene activities and molecular
composition won't ever tell us what kind of physical system we are dealing
with and what its properties are. Since evolution is all about organisms
and how they change, if we don't understand how they are generated and what
kinds of transformation they undergo, we are going to have trouble understanding
what evolution is about.
This is where natural selection is usually brought in as an explanatory
principle. Why do plants have leaves? Because leaves aided survival and
reproduction. Why do vertebrates have limbs? Because limbs helped them to
get around in the world, and thus to survive and reproduce. The issues here
are about stability and instability: the capacity of a species to persist
in particular environments and the extinctions that follow when perturbations
exceed its capacity for adaptive response. In the study of dynamic systems,
stability is described in terms of attractors, the states into which systems
settle such as the point at the bottom of the bowl where a marble, released
at the top of the bowl, eventually stops ; or the steady state of a population
in which birth and death rates balance.
Just how dynamic these can be is demonstrated by
the strange attractors of systems governed
by deterministic chaos. These attractors are described
as "strange" because the motion never repeats itself but wanders in a restricted
domain. So the appropriate concept for studying the capacity of species to
survive is dynamic stability, and this is what natural selection is about.
But dynamic stability does not explain the form of organisms. True, the forms
of existing species must have been sufficient for their survival. But this
statement gives us virtually no insight into the particular structure and
behaviour of organisms such as the specific pattern of leaves on a plant
or the flight of the bumble bee.
What 20th century biology does give us - and this point is particularly clearly
described in Dawkins' books - is an extremely successful and important theory
of biological inheritance and the molecular composition or organisms. The
result is that we can now say a great deal about gene frequencies and their
changes in populations, about the remarkable ways in which genomes behave
both as the vehicles of inheritance and as the coding and control systems
for molecular syntheses, and many other aspects of organisms and their evolution.
But what is missing from the dazzling array of discoveries that constitute
20th century biology is any fundamental theory about organisms as self-
generating entities, and the principles of their organisation as dynamic
systems of a particular kind. Self-reproduction is now understood at the
level of DNA replication, and a magnificent achievement this is. But DNA
replication is in no sense equivalent to the reproduction of organisms. In
a test-tube, a DNA or an RNA replication system gets simpler and simpler,
replicating faster and faster until it reaches a limit defined by the
coding sequences required for replication
itself (Spiegelman, 1967). This in vitro demonstration illustrates
precisely the properties of dynamic stability discussed earlier: the attractor
for this system is a stable state in which the simplest, fastest replicators
have taken over and displaced all slower replicators from the reaction
system.
But, organisms can go the opposite way : they can become much more complex.
As we often see in evolution, such elaborations cannot be explained by the
properties of DNA. Furthermore, the genomes of higher organisms cannot be
accurately replicated except in the context of the reproducing organism.
So biological reproduction is an organismic property that cannot be reduced
to the behaviour of replicators.
How is a biological theory of the form of organisms to be developed? Such
a theory will be developed in exactly the same way as in any other science.
The properties of organisms are studied both experimentally and theoretically,
developing a generative theory that describes how their morphologies and
behaviour patterns arise and what sorts of transformations they can undergo.
The insights that have been provided by modern biology are extremely important
to such an endeavour, but these are used within the context of a dynamic
analysis of the three dimensional forms and patterns that are generated by
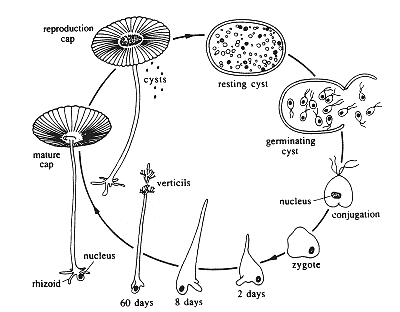
the organism during its life cycle. To illustrate this point, consider a
simple life cycle such as that of the unicellular green algae,
Acetabularia, shown in Figure 1. What we want to know is how and why
the organism goes through the sequence of shape changes that characterize
this particular life cycle, and what other shapes may become possible when
either the genes or the environment are changed. To investigate the origins
of organismic form, we must develop an explicit theory that describes the
three- dimensional transformations of the organism in terms of a field theory
(Goodwin and Briere, 1992) that takes account of the spatial patterns of
the system and their changes in time. Developing organisms are described
as morphogenetic fields, and in constructing the theory, we use detailed
information about the molecular organization of the cytoplasm and the cell
wall that underlies the capacity of the organism to grow and change its shape.
Equations are derived to describe the mechanical
properties of the cytoplasm that arise from the interactions between the
cytoskeleton and calcium. These show how spatial
patterns of strain (stretch or compression) and of cytoplasmic calcium
concentration can arise spontaneously , initiating local changes of shape
and differential growth in the developing organism. These patterns are studied
in three dimensions on a computer graphics display using what is called a
finite element description of the system.
The type of question that can then be asked is how and why the organism goes
through the sequence of shape changes that characterize this particular life
cycle, and what other shapes may be possible by changing either the genes
or the environment. The issues raised here about organisms are well illustrated
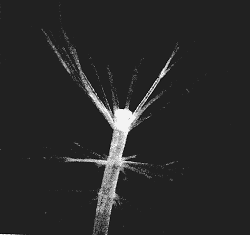
by the structures called whorls that are produced during the development
of Acetabularia. These can be seen as the rings of finely-branching
filaments called laterals (rather like primitive leaves), shown in Figure
2, which is a cell with three such rings or whorls of laterals and the beginning
of a cap. In the adult, the whorls are absent - the laterals have all dropped
off Why are these structures made when they apparently serve no purpose?
Now we can never know precisely what function particular structures may play
in organisms. However, in our experimental studies we have shown that
Acetabularia can grow perfectly well and can even make caps and reproduce
without ever making whorls. So the whorls don't seem to be of much value.
Why, then, are they made? There may be a very simple answer. They are made
because the dynamic organization of the system typically (or generically
, to use the mathematical term) produces whorls, just as the dynamic organization
of liquids typically (generically) results in spiral flow down drains.
|
| Fig1.The Life cycle of Acetabularia acetabulum,showing the different stages of morphogenesis including the production of the transient verticils (whorls of laterals). |
This suggestion comes from modelling studies. We observed
that once the parameters in the equations describing growth and morphogenesis
were set so that localized tip formation could occur, subsequent changes
of form happened spontaneously giving a sequence of whorls. Genes can be
regarded as determining parameter values, so they play an important role
in setting up the dynamics. But then the organization of the system takes
over and leads to whorls without further changes of parameter, morphogenesis
apparently leading to a generic form in what Gould ( 1991 ) calls morphospace,
the space of potential forms. Whorls of order to which Acetabularia
belongs, the Dasycladales, have been around for at least 570 million years.
In most species the whorled laterals act as gametophores, where gametes are
produced. Only in the Acetabulariaceae are they supplanted by caps
as the gametophores, though whorls continue to be made. What this suggests
is that such high-level taxonomic characters are robust consequences of
morphogenesis typical of a particular type of organism, due to their basic
organisation (described in a particular type of morphogenetic field). So,
the Acetabulariaceae have whorls not because they are useful (adapted)
but because they are typical of the organization of the order to which they
belong. Genetic differences can result in variations on this theme; but beyond
a certain range of genetic change the whole morphogenetic sequence changes
and a different generic form arises. This then defines another taxonomic
group. Evolution at this level is transformation between generic forms.
|
| Fig2. An algae with three whorls and the beginnings of a cap. |
So, my conclusion is that organisms are not survival machines constructed
by selfish genes, machines whose behaviour is dictated by natural selection.
They are self- generating wholes, complex systems that obey principles of
dynamic order described by field equations. These dynamic forms use their
DNA to specify parameters of the process so the parameters replicate along
with the life cycle and result in repeatability of the morphogenetic process.
The whole reproductive cycle must be stable if the species is to survive.
So we accommodate all of the basic properties of genes and the exigencies
of natural selection, but within the context of a theory of the organism
and its , life cycle as the fundamental entity in biology. As defined, organisms
are not machines, they are not reducible to genes and gene products, and
their properties are not explained by natural selection. They must be understood
as self-generating, reproducing structures of a distinct kind whose
transformations define the range of living forms available for evolution.
RICHARD DAWKINS : ORGANISMS MAY BE WHOLES BUT ONLY GENES ARE
REPLICATORS
What is a motor car? A motor car is a means of personal transportation.
No it isn't; a motor car is a metal box with padded seats, a wheel at each
corner and an internal combustion engine.
No no, you're both wrong; a motor car is the product of a factory where it
is assembled from parts separately manufactured, usually on a production
line, to a design drawn on a drawing board.
Nonsense, a motor car isn't any of those things. A motor car is a status
symbol ... the curse of the age ... a boon to the country doctor ... the
single greatest contributor to atmospheric pollution ... where most American
teenagers lose their virginity ... a lethal weapon in the wrong hands ...
a horseless carriage, the twentieth century descendant of the coach and four
... a death-trap on ice ....
I hope it is clear that the only thing seriously wrong with each of these
answers is the No , that begins it. A motor car can be more than one thing
at a time. In particular, we can recognize my first three answers as the
functional answer, the structural answer and the developmental answer. All
three are true. All three are important. None can, in any sense whatsoever,
be regarded as a substitute for either of the others. They are three distinct
truths, simultaneously true about the same object.
The same applies to the question, "What is an organism?" Goodwin may be right
that organisms are "self-generating wholes, complex systems that obey principles
of dynamic order described by field equations." But I am simultaneously right
that organisms are survival machines constructed by selfish genes that have
weathered natural selection. Brian Goodwin is an embryologist. The questions
he asks are development questions. These are important questions, and his
answers are clever and possibly correct answers. But there are other questions
that are simultaneously interesting and worthwhile- If Goodwin's answer to
his developmental question is right, this may imply that other answers to
the same question are wrong. But it does not imply that answers to different
questions are wrong. For the sake of argument, let' s accept Goodwin' s
characterization of organisms as self- generating wholes governed by field
equations. Suppose Goodwin is right that the organism develops as "a functional
and a structural unity, in which the parts exist for and by means of one
another" as opposed to a system, like a car in which the parts "pre-exist
before being assembled into a functioning unity." Well and good, but, as
we shall see, some of us might feel the need to ask why natural selection
has favoured the one kind of embryology over the other. The explanation will
be a Darwinian explanation. According to our modem understanding of Darwinism,
that means an explanation in which genes fostering the one embryology survive
better than genes fostering the other.
But what, in my world view , is so special about genes anyway? If Goodwin
is right that they play only a minor role in determining the forms of organisms,
minor in comparison to the principles of dynamic order, why thrust genes
to centre stage? The reason is that genes have a unique role to play in
functional explanations. Genes do two quite different things. They affect
embryonic development within an individual, and they replicate themselves
down the lineages of different individuals. Replicating entities are sifted
by natural selection and, if they are successful in the sifting, they potentially
go on replicating forever. Given spontaneous variation due to mutation, the
world becomes full of successful genetic replicators at the expense of
unsuccessful ones. But what makes some genetic replicators successful and
others not? What do the winners have that the losers lack? This is where
the other main activity of genes comes in, their embryological influence.
Genes are successful, or unsuccessful, because of their effects on embryonic
development. Unlike the blueprints of a car, genes have to ride inside the
vehicles whose development they influence. They have to live with their mistakes.
In their mistakes, even! If the vehicle falls over a cliff, so do
they . If the vehicle is devoured by a leopard, so are they. As the generations
go by, therefore, the world automatically becomes full of copies of genes
whose products don't fall over cliffs and are not eaten by leopards. Every
creature born is descended from an unbroken line of successful ancestors,
and they inherit what it takes to be successful.
The business of surviving to reproduce becomes extremely complicated, especially
with enemies simultaneously evolving ever more ingenious tricks of predation,
parasitism, escape. Not surprisingly, in the course of these arms races,
genes have evolved ever more complicated mechanisms for staying alive and
reproducing: survival machines.
And why do we say that these survival machines work "for" the organism's
genes, rather than for, say, its teeth or ankles? After all, if the organism's
ankles fail and it stumbles in the path of a leopard, the ankles perish along
with the genes. Ankles, too, have to live with their mistakes. Why isn't
the organism a survival machine for ankles? The answer is simple. Genes are
replicators. Ankles are not. You may be tempted to think that ankles are
replicators, because .they reappear in every generation. But your ankles
are not replicas of your parents' ankles. This is not only because you have
two parents. It is also because changes in ankles, unlike changes in genes
affecting ankles, are not inherited. If they were, babies would be born with
ready-sprained ankles covered with varicose veins. But
"sprain" a gene, for example with X-rays
or a mutagenic chemical, and the baby really will inherit the altered form.
What is special about genes in evolutionary explanations is that, amid the
welter of complexity that enters into the development of a body, genes and
only genes (setting aside the inheritance of cultural tradition) have the
potential for being replicated down the generations with errors that are
themselves replicated. That is why bodies are survival machines for genes.
Brian Goodwin does have an interesting point to make. Just as the whirlpool
in the bath-waste settles down into one of two stable spiral patterns, and
the only freedom for variation is whether it is clockwise or anticlockwise,
so body form may be similarly constrained. There may be a limited set of
alternative stable states, and genetic mutation may be free only to kick
embryonic development from one to another of them. Everything that is interesting
and beautiful about a whirlpool comes from the physics of vortices, and all
that your foot can do when you get out of the bath is nudge it towards the
clockwise or the anticlockwise mode. In the same way, it might be that all
that is interesting and beautiful about living organisms comes from field
equations, not from differences between genes, which can only nudge the
organism's form into the animate equivalent of a clockwise or anticlockwise
spiral.
But Goodwin, like his predecessor D'Arcy Thompson ( 1942), is wrong to think
that his views are in any way anti-Darwinian. Even if Goodwin's vision were
true in its extreme form it would still not undermine the statement that
adaptations, to the extent that they are for the benefit of something, must
be for the benefit of self- replicating entities-genes. Field equations,
unlike genes, do not exhibit heredity. Whirlpools don't beget lineages of
daughter whirlpools that inherit their particular attributes- If they did,
they would be true replicators and might well engender an evolution of their
own- In any case,I am pretty sceptical of Goodwinian embryology in its extreme
form. This is because living organisms are so beautifully and elaborately
"designed." No doubt embryology does impose some constraints on what mutation
and selection are allowed to get away with: doubtless there are some "attractors
in morphospace." But it seems intuitively clear that organisms must be miles
away from the "whirlpool" end of the spectrum, simply because they are so
functionally elegant. Organisms are finely tuned to survive, honed to the
last detail. If they were subject to the sort of constraints that hem whirlpools
in, organisms wouldn't be so exquisitely good at what they do. Just open
any page of, say, Cott's (1940) Adaptive Coloration or Wickler's (1968)
Mimicry, Hansell's (1984) Animal Architecture or, for that
matter, Darwin's ( 1882) Orchids, and ask yourself if this sort of
thing could be achieved by highly constrained, Goodwinian embryology.
But now here is a more constructive overture towards Brian Goodwin. I have
represented "whirlpool embryology" as a constrained embryology. Let me change
the metaphor to show what a difference it makes: let's consider "kaleidoscopic
embryology". Because of the arrangement of mirrors or prisms in a kaleidoscope,
random heaps of coloured chips cannot help looking pretty. This is thanks
to the constraint of radial symmetry. Random mutations-knocks on the barrel
that change the positions of the chips-set off intricate and elegant changes
at all corners of the radically symmetrical pattern. We can say that the
shapes are constrained by the mirrors to be radically symmetrical, but
"constrained" sounds too negative. We can also regard the mirrors as positively
designed to make the patterns more elaborate and captivating, perhaps via
symmetry in various planes (in which case the metaphor of the kaleidoscopic
mirror is especially apt), or perhaps via segmentation or other kinds of
controlled repetition (Dawkins, in preparation).
In my essay The Evolution of Evolvability (Dawkins 1989), I have suggested
that natural selection has not only favoured genes that make organisms good
at surviving and reproducing. It has also favoured embryologies that are
, "good at evolving." To suggest that mutations could anticipate future changes
in the environment is rightly regarded as heresy (Williams, 1966). But it
is not out of the question that some form of selection could pick out those
embryologies that - perhaps through being kaleidoscopic - have proved
evolutionarily fruitful. It is still right to regard each new mutation as
random with respect to function. But mutations have to work by altering the
existing embryology. And not all embryologies are equally pregnant with
evolutionary responsiveness, equally luxuriant in mutational possibility.
Perhaps natural selection has fostered "kaleidoscopic" embryologies because
of their evolutionary plenitude. If organisms develop "as self-generating
wholes, complex systems that obey principles of dynamic order described by
field equations," it is because natural selection has favoured this kind
of embryology in the past. Perhaps it has done so not just because such
embryologies make organisms that are good at surviving and reproducing. Perhaps,
I am suggesting, this kind of embryology is favoured because it consistently
unleashes cascades of potent mutations-it is good at evolving. Instead of
belittling Darwinism, I think that Brian Goodwin should see his version of
embryonic development as one of Darwinism's greatest achievements and the
key to the blossoming of further great achievements.
BRIAN GOODWIN: GENES MAY BE REPLICATORS BUT IT IS ORGANISMS THAT
EVOLVE
Richard Dawkins' modified position is so reasonable that I am tempted to
say that we are agreed in principle but disagree over details and emphasis.
A transformed Darwinism in which organisms are recognized as dynamic wholes
with distinctive principles of organization that underlie their capacity
to develop, reproduce, and evolve is an enormous step away from the reductionism
of selfish genes and their survival in gangs that cooperate just to leave
more copies of themselves. It brings biology into line with mainstream
contemporary studies of complex systems such as
fractals and
chaos in which it is recognized that, underlying
what appears to be kaleidoscopic variety of form and behavior there are
principles of order and organization that make the diversity intelligible.
This is the position I am arguing for: that organisms, despite their remarkable
diversity of form, belong to a class of organized dynamical system with deep
principles of order that cannot be understood in terms simply of gene activities,
nor be explained as the result of natural selection. I have no desire to
ignore or belittle the importance of genes, but they have to be understood
in context, which is the dynamic form we call an organism. Gene activities
define molecular composition, which specifies parameters such as kinetic
constants of reactions via enzymes, binding constants of calcium- regulating
proteins, and the viscous and elastic
constants of cytoplasm; natural selection relates to the stability of life
cycles. Important as these factors are, even together they are not sufficient
to explain the properties of organisms and their diversity as members of
the particular class of dynamical system we call living.
There are ways of studying and classifying the members of this set, which
define different species, by experimental and theoretical methods. I described
one approach briefly in relation to experimental, mathematical and computer
modelling of the species Acetabularia acetabulum, giving an explanation
for structures that are not easily explained by natural selection but are
readily understood in terms of the developmental dynamics of this organism.
If Dawkins' "kaleidoscopic embryology" means acknowledging the study of whole
organisms and their transformations (developmental and evolutionary) as a
basic and fundamental biological enterprise, with genes specifying essential
factors (parameter values and other aspects of the dynamics) of life cycles
but not themselves sufficient to explain orgasmic form and behavior, then
we're agreed in principle. However, let' s not fudge the issue: this description
of organisms inverts the emphasis from that in Neo-Darwinism, which focuses
on inheritance and natural selection and largely ignores the generative
principles of organisms and their life cycles (dynamics). In the biology
I'm proposing, nothing of value in Neo-Darwinism is lost, but whole organisms
as life cycles are the irreducible entities that evolve, with genes and natural
selection as aspects of these dynamic cycles and their stability in particular
environments. For a comprehensive evolutionary theory it is necessary to
go further than this, into the dynamics of ecosystems, but these should be
constructed also out of interacting organisms (life cycles) and their
environments, not out of genes.
Richard Dawkins' final remarks make it clear that he stops short of an
organism-based biology, continuing to see natural selection as the explanatory
principle of evolution, not the dynamics of organisms and their interactions.
Natural selection has favoured embryologies that are , "good at evolving,"
he says. But this is inevitable: those systems that generate more stable
dynamic processes will be more numerous, whether in biology, chemistry, or
physics. It is just the stability argument, used at another level: a system
with the potential of producing many different stable states rather than
just a few will have more dynamical "offspring" that "survive." Hence it
will be better at evolving. But we need to describe the actual dynamics of
such biological systems, the developmental dynamics of life cycles and other
aspects of biological organization, to understand their intrinsic order and
what organizational properties underlie differences of potential for evolution.
A recent and very interesting conjecture is that there is a dynamic attractor
in complex systems "at the edge of chaos," which is the "best" place for
evolving systems to be (Kauffman, 1993).
There is a problem in Dawkins' argument that I must consider, relating to
adaptation- He sees adaptation as the result of the selection of genes by
nature. The problem is that how we think about adaptation is also a result
of selection, the selection of examples of adaptation by Darwinian biologists.
While there are plenty of instances of remarkable adaptations, from the striking
mimicry by one butterfly species of another's
wing pigment patterns to the perfection of the eyes of cats and owls, there
are as many non-adaptations that seem to cause no survival problems. I have
already mentioned the whorls on Acetabularia. In addition, there are
wasps of several families that spend most of their time under water, using
their wings to swim. These wasps look just like their land-dwelling relatives.
No adaptations here. [That can't be true.In order to breathe underwater
there must be some adaptation from an air breathing variety. -LB]
Similarly, flowering plants have taken over most of the earth, equipped with
superior morphological and reproductive mechanisms, yet ferns, horsetails,
clubmosses, and their relatives with the relatively primitive structures
of 200-million-year-old ancestors "compete" successfully.
[Such "competition" is misunderstood if niches are accounted for.If there
is no threat to a niche or an organism develops a good strategy in the first
instance there is little need to update it.Certain species of crab still
look similar to those though to have existed in prehistory.The Nautilus seems
unchanged when compared to fossil versions.If an organism has found a quiet
cul-de-sac,there is little requirement for change.Competition happens where
there is dispute over a niche.-LB]
It looks more like anything goes that works well enough, rather than constant
fine-tuning of species to habitats, the expectation of competition and
adaptations whereby species are constantly climbing hills in fitness landscapes
to out-do their competitors.
Finally some of the Neo-Darwinists' favourite examples can be explained in
developmental terms. "Adaptations" such as butterfly mimicry can most readily
be explained by constraints on development, which have the consequence that
only a limited number of pigment
patterns is possible. So it is not difficult for one species to develop
a pattern similar to that of another. The phenomenon of pseudomimicry greatly
strengthens this view . In this case, species that inhabit totally different
regions-such as Puerto Rico and Indonesia- nevertheless have similar pigment
patterns. What advantage could this possibly give them? Understanding the
principles according to which pigment patterns develop on wings as studied
by F. Nijhout (1991), provides ways of explaining why both types of mimicry
are not only possible, but probable. Again, we have to look to the organism
for answers to all of these questions. When we have done this systematically
, biology is going to look like a rather different science from its present
form.
 |
Simplified Tree of Life "Life on Earth" D.Attenborough (p310) Click to Expand |